Recombinant Human BAG family molecular chaperone regulator 3 (BAG3)
-
中文名稱:Recombinant Human BAG family molecular chaperone regulator 3(BAG3)
-
貨號(hào):CSB-YP002531HU
-
規(guī)格:
-
來(lái)源:Yeast
-
其他:
-
中文名稱:Recombinant Human BAG family molecular chaperone regulator 3(BAG3)
-
貨號(hào):CSB-EP002531HU
-
規(guī)格:
-
來(lái)源:E.coli
-
其他:
-
中文名稱:Recombinant Human BAG family molecular chaperone regulator 3(BAG3)
-
貨號(hào):CSB-EP002531HU-B
-
規(guī)格:
-
來(lái)源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Human BAG family molecular chaperone regulator 3(BAG3)
-
貨號(hào):CSB-BP002531HU
-
規(guī)格:
-
來(lái)源:Baculovirus
-
其他:
-
中文名稱:Recombinant Human BAG family molecular chaperone regulator 3(BAG3)
-
貨號(hào):CSB-MP002531HU
-
規(guī)格:
-
來(lái)源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:BAG 3; BAG family molecular chaperone regulator 3; BAG-3; Bag3; BAG3_HUMAN; Bcl 2 binding protein; Bcl-2-associated athanogene 3; Bcl-2-binding protein Bis; BCL2 associated athanogene 3; BCL2 binding athanogene 3; BIS; CAIR 1; Docking protein CAIR 1; Docking protein CAIR-1; MFM6
-
種屬:Homo sapiens (Human)
-
蛋白長(zhǎng)度:Full Length of Mature Protein
-
表達(dá)區(qū)域:2-575
-
氨基酸序列SAATHSPMM QVASGNGDRD PLPPGWEIKI DPQTGWPFFV DHNSRTTTWN DPRVPSEGPK ETPSSANGPS REGSRLPPAR EGHPVYPQLR PGYIPIPVLH EGAENRQVHP FHVYPQPGMQ RFRTEAAAAA PQRSQSPLRG MPETTQPDKQ CGQVAAAAAA QPPASHGPER SQSPAASDCS SSSSSASLPS SGRSSLGSHQ LPRGYISIPV IHEQNVTRPA AQPSFHQAQK THYPAQQGEY QTHQPVYHKI QGDDWEPRPL RAASPFRSSV QGASSREGSP ARSSTPLHSP SPIRVHTVVD RPQQPMTHRE TAPVSQPENK PESKPGPVGP ELPPGHIPIQ VIRKEVDSKP VSQKPPPPSE KVEVKVPPAP VPCPPPSPGP SAVPSSPKSV ATEERAAPST APAEATPPKP GEAEAPPKHP GVLKVEAILE KVQGLEQAVD NFEGKKTDKK YLMIEEYLTK ELLALDSVDP EGRADVRQAR RDGVRKVQTI LEKLEQKAID VPGQVQVYEL QPSNLEADQP LQAIMEMGAV AADKGKKNAG NAEDPHTETQ QPEATAAATS NPSSMTDTPG NPAAP
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Co-chaperone for HSP70 and HSC70 chaperone proteins. Acts as a nucleotide-exchange factor (NEF) promoting the release of ADP from the HSP70 and HSC70 proteins thereby triggering client/substrate protein release. Nucleotide release is mediated via its binding to the nucleotide-binding domain (NBD) of HSPA8/HSC70 where as the substrate release is mediated via its binding to the substrate-binding domain (SBD) of HSPA8/HSC70. Has anti-apoptotic activity. Plays a role in the HSF1 nucleocytoplasmic transport.
-
基因功能參考文獻(xiàn):
- There was no association of SNPs in ADRB1, GRK5 and BAG3 genes with Takotsubo cardiomyopathy. PMID: 29514624
- These findings provide a molecular basis for understanding of BAG3dependent cell proliferation and survival from the aspect of alteration of gene expression. PMID: 30106105
- BAG3 can modulate the levels, localization or activity of its partner proteins, thereby regulating major cell pathways and functions, including apoptosis, autophagy, mechanotransduction, cytoskeleton organisation, motility. PMID: 28864347
- Genetic and pharmacological interference with BAG3 is capable to resensitize TNBC cells to treatment. PMID: 29462756
- The HSF1-BAG3-Mcl-1 signal axis is critical for protection of mutant KRAS colon cancer cells from AUY922-induced apoptosis. PMID: 29068469
- The Hsp70-Bag3 complex therefore functions as an important signaling node that senses proteotoxicity and triggers multiple pathways that control cell physiology, including activation of protein aggregation. PMID: 29987014
- study revealed oncogenic roles of BAG3 in chondrosarcoma and provided mechanisms that the BAG3-modulated the expression of RUNX2 through upregulation of beta-catenin. PMID: 29484408
- BAG3 interacted with CXCR4 mRNA and promoted its expression via its coding and 3'-untranslational regions. PMID: 28703799
- Silencing of HSPB8 markedly decreased the mitotic levels of BAG3 in HeLa cells, supporting its crucial role in BAG3 mitotic functions. The results support a role for the HSPB8-BAG3 chaperone complex in quality control of actin-based structure dynamics that are put under high tension, notably during cell cytokinesis. PMID: 28275944
- Our findings suggest that high levels of BIS expression might confer stem-cell-like properties on cancer cells through STAT3 stabilization PMID: 27145367
- modulation of proteostasis is a distinct biological function of sAPPalpha and does not require surface-bound holo-APP. PMID: 26526841
- higher levels of BAG3 were observed in hypertensive patients compared to healthy controls, and even higher levels in hypertensive diabetic patients compared to healthy subjects. PMID: 28696030
- BAG3 directly stabilizes hexokinase 2 mRNA and promotes aerobic glycolysis in pancreatic cancer cells. PMID: 29114069
- These results indicated that at least some oncogenic functions of BAG3 were mediated through posttranscriptional regulation of Skp2 via antagonizing suppressive action of miR-21-5p in ovarian cancer cells. PMID: 28624440
- Familial suffering of Dilated cardiomyopathy and carrying a heterozygous large deletion in the BAG3 gene.This gene encodes BCL2-associated athanogene 3 protein. PMID: 27736720
- BAG3 mutations are associated with DCM phenotypes. BAG3 should be added to cardiomyopathy gene panels for screening of DCM patients, and patients previously considered gene elusive should undergo sequencing of the BAG3 gene. PMID: 28211974
- The spatial regulation of mTORC1 exerted by BAG3 apparently provides the basis for a simultaneous induction of autophagy and protein synthesis to maintain the proteome under mechanical strain. PMID: 27756573
- This study therefore identifies both BAG3 reduction and autophagy promotion as potential therapies for FLNC(W2710X) myofibrillar myopathy, and identifies protein insufficiency due to sequestration, compounded by impaired autophagy, as the cause. PMID: 26969713
- variants in TNNT2 and BAG3 are associated with a high propensity to life-threatening cardiomyopathy presenting from childhood and young adulthood. PMID: 28669108
- It has been demonstrated that HSPB8-BAG3-HSP70 ensures the functionality of stress granules and restores proteostasis by targeting defective ribosomal products for degradation. PMID: 27570075
- Study report p.H243Tfr*64_BAG3 as a novel pathogenic variation responsible for familial dilated cardiomyopathy. This variation correlates with a more severe phenotype of the disease, mainly in younger individuals. PMID: 27391596
- The authors propose that the chaperone-mediated autophagy function of BAG3 represents a specific host defense strategy to counteract the function of VP40 in promoting efficient egress and spread of virus particles. PMID: 28076420
- BAG3 bound to Hsp70 at the same time as Hsp22, Hsp27, or alphaB-crystallin, suggesting that it might physically bring the chaperone families together into a complex. PMID: 27884606
- These results suggest that Bag1 and Bag3 control the stability of the Hsc70-client complex using at least two distinct protein-protein contacts, providing a previously under-appreciated layer of molecular regulation in the human Hsc70 system. PMID: 27474739
- we report the first mammalian model of a single amino acid mutation of BAG3 (P209L) in exon 3 of Bag3 associated with the development of muscle disease with left ventricular dysfunction and heart failure PMID: 27321750
- High BAG3 expression Correlates with Sebaceous Gland Carcinoma of the Eyelid. PMID: 28373462
- These results indicate a possible role for BAG3 protein in the maintenance of cell survival in endometrioid endometrial cancer. PMID: 27414463
- By showing transcription factor HSF1 activation, we demonstrated that HCA induces the expression of BAG3 through HSF1 activation. More importantly, knockdown of BAG3 expression using siRNA largely inhibited HCA-induced apoptosis, suggesting that BAG3 is actively involved in HCA-induced cancer cell death PMID: 27922674
- These results provide further insight into the molecular mechanisms involved in the enhancement of hyperthermia sensitivity by the silencing of BAG3 in human oral squamous cell carcinoma cells. PMID: 27245201
- high expression of BAG3 was detected in a majority of medulloblastoma tissues and predicted poor outcome for medulloblastoma patients. PMID: 27456361
- in the present study, we demonstrated that BAG3 overexpression plays a critical role in cell proliferation, migration, and invasion of colorectal cancer. Our data suggests targeted inhibition of BAG3 may be useful for patients with colorectal cancer PMID: 26577854
- BAG3 maintains the basal amount of LC3B protein by controlling the translation of its mRNA in HeLa and HEK293 cells. PMID: 26654586
- Data indicate a tumor suppressor-like function of Bcl-2 associated athanogene 3 (BAG3) via direct interaction with glucose 6 phosphate dehydrogenase (G6PD) in hepatocellular carcinomas (HCCs) at the cellular level. PMID: 26621836
- BAG3 is associated with Z-disc maintenance. PMID: 26545904
- BAG3 is a suitable target for combined therapies aimed at synergistically inducing apoptosis in bladder cancer. PMID: 26100943
- Bis expression was higher in squamous cell carcinoma than in adenocarcinoma in Lung Cancer. PMID: 25968616
- BAG3 promotes pancreatic ductal adenocarcinoma growth by activating stromal macrophages. PMID: 26522614
- BAG3-mediated miRNA let-7g and let-7i inhibit proliferation and enhance apoptosis of human esophageal carcinoma cells by targeting the drug transporter ABCC10 and modulates cisplatin resistance. PMID: 26655271
- This newly described ERa-mediated and estrogen response element (ERE)-independent non-canonical autophagy pathway, which involves the function of BAG3 and provides stress resistance in our model systems. PMID: 26158518
- BAG3 protein loci is involved in the pathophysiology of systolic heart failure. PMID: 25889438
- MiR-143 enhanced the tumor suppressive effect of shikonin partly through the regulation of BAG3 in glioblastoma stem cells. PMID: 26541455
- The Hsp70-Bag3 interaction may be a promising, new target for anticancer therapy. PMID: 25564440
- BAG3 mutations should be considered even in cases with a mild phenotype or an adult onset PMID: 25208129
- The stress response protein BAG3 may be prognostic for death in patients with acutely decompensated heart failure. PMID: 25753466
- BAG-3 expression correlated with increased HSP70 expression in a subset of systemic T cell lymphoma cases co-expressing the CD30 antigen. PMID: 24492285
- To examine the relationships among BAG3, miR-29b and MMP2 in endometrioid adenocarcinoma cells. PMID: 25738313
- Upon glucose stimulation BAG3 is phosphorylated by FAK and dissociates from SNAP-25 allowing the formation of the SNARE complex, destabilization of the F-actin network and insulin release. PMID: 25766323
- Studies indicate that heat shock protein Hsp70 (Hsp70) regulates multiple pathways in cancer cells via interaction with Bcl2-associated athanogene 3 protein (BAG3) co-chaperone. PMID: 25347739
- BAG3 plays a key role in the processing of the nucleocytoplasmic shuttling of HSF1 upon heat stress PMID: 26159920
- HIV-1 Tat protein is able to stimulate autophagy through increasing BAG3 levels in glial cells. PMID: 25483098
顯示更多
收起更多
-
相關(guān)疾病:Myopathy, myofibrillar, 6 (MFM6); Cardiomyopathy, dilated 1HH (CMD1HH)
-
亞細(xì)胞定位:Nucleus. Cytoplasm.
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
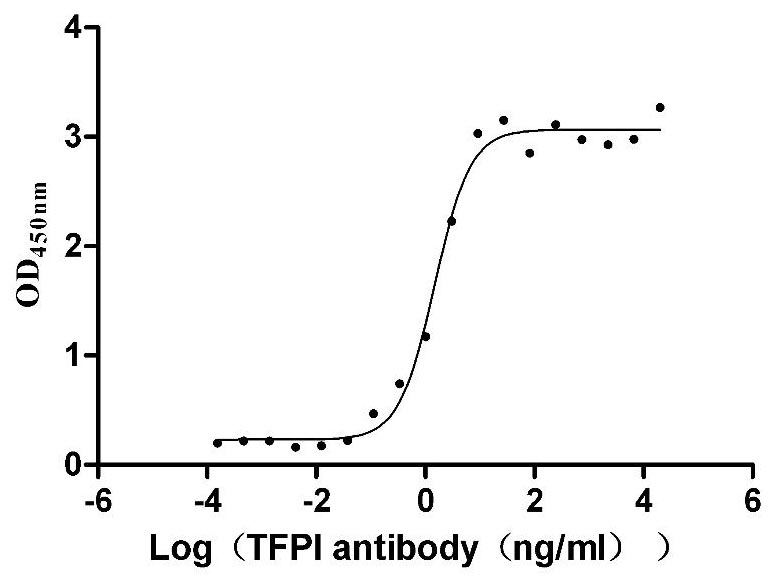
Recombinant Human Tissue factor pathway inhibitor (TFPI), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
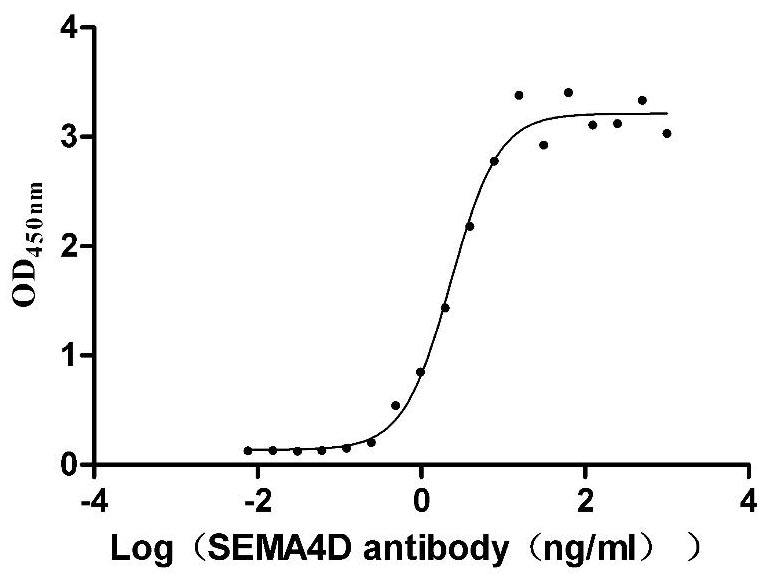
Recombinant Macaca mulatta Semaphorin-4D isoform 1 (SEMA4D), partial (Active)
Express system: Mammalian cell
Species: Macaca mulatta (Rhesus macaque)
-
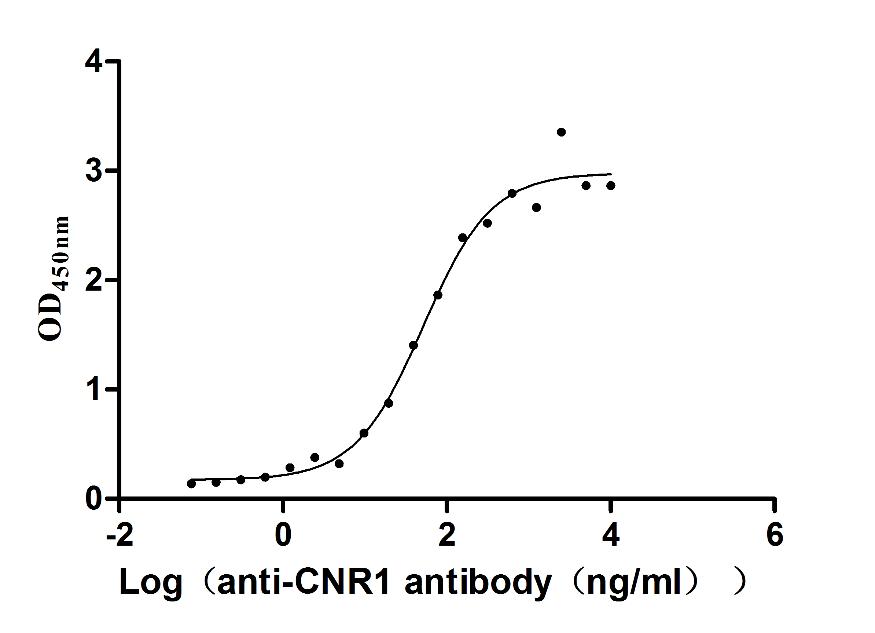
Recombinant Human Cannabinoid receptor 1 (CNR1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
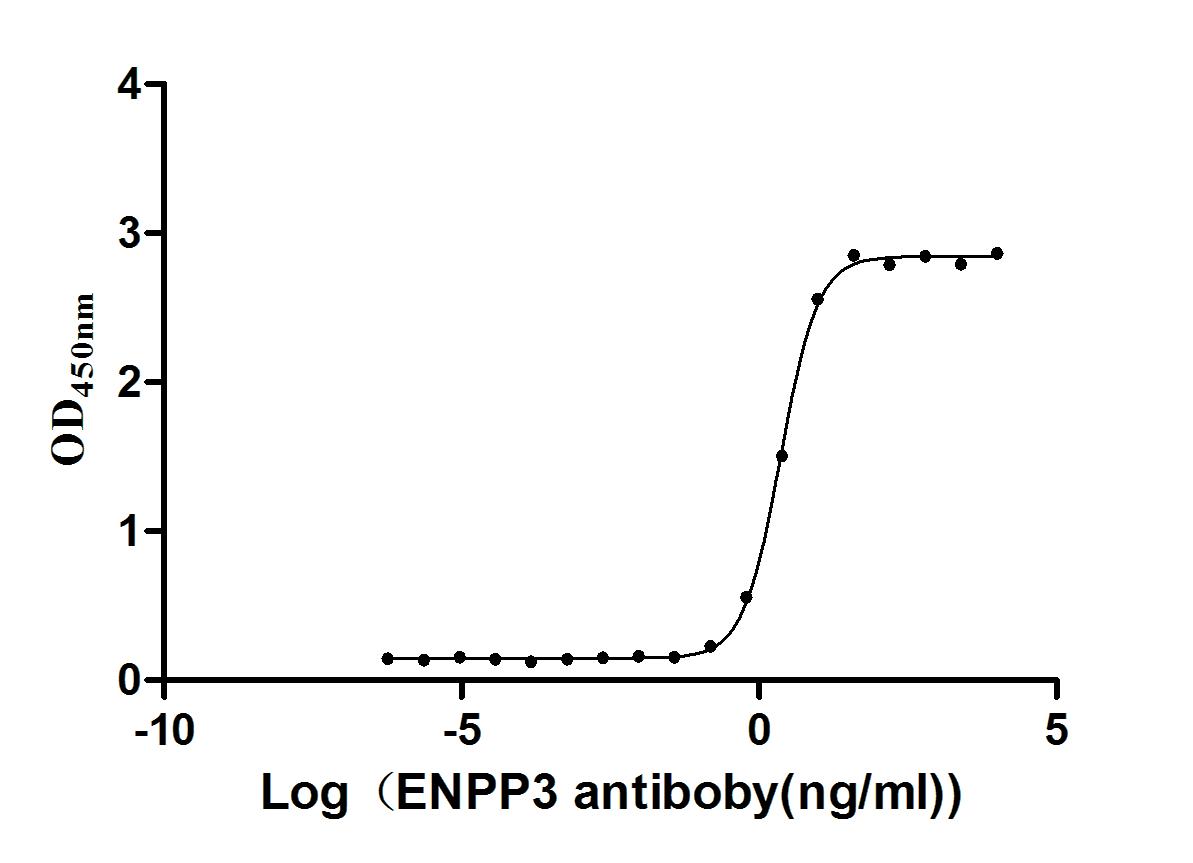
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Dipeptidase 3(DPEP3), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)





")
")






